
Viele Pflanze können sich vegetativ vermehren. Das bedeutet vervielfältigen, ohne Keimzellen zu bilden. Dabei entsteht jedoch nur eine vielzahl von Individuen, jedoch keine veränderung im genetischen Material. Die Nachkommen sind sozusagen Klone.
Beispiele für Vegetative Vermehrung sind sogenannte Stecklinge, also Sprossabschnitte die in die Erde gesteckt wieder zu vollständigen Pflanzen auswachsen, Brutkörperchen wie Brutblätter bei Scharbockskraut und Poa alpina, Tochterzwiebeln, Ausläufer wie bei der Erdbeere, Knollen wie bei der Kartoffel, Blätter bei Crassulaceen und in Zellkulturen können sogar einzelne Zellen zur vegetativen Vermehrung genutzt werden.
Generationswechsel
Um eine Vervielfältigung der Individuenzahl und eine vielfalt von genetischen Variationen zu gewährleisten machen Pflanzen einen sogenannten Generationswechsel.
Ein Generationswechsel ist dabei als die regelmäßige Abfolge von Generationen definiert, welche mit einem Typ Fortpflanzungszellen beginnen und mit einem anderen Typ enden.
Sporen -> Gametophyt -> Gameten
(Gameten) Zygote -> Sporophyt -> Sporen
Die meisten Landfarne sind dabei Isospor, das heißt der Gametophyt bildet sowohl die Eizelle, als auch die Spermatozoiden welche zusammen finden und gemeinsam die Zygote bilden.
Die Wasserfarne, Moosfarne und Samenpflanze dagegen sind heterospor, das heißt ein Individuum bildet die Eizelle und ein anderes die Spermatozoiden, welche gemeinsam, also mit dem gemischten Genmaterial von 2 Individuen den Gametophyt bilden.
Farne, als auch Samenpflanzen machen dabei einen heterophasischen-heteromorphen Generationswechsel. Heterophasisch heißt hierbei, dass eine Generation diploid ist und die andere haploid. Heteromorph bezeichnet dagegen nur das unterschiedliche Aussehen der beiden Generationen.
Bei Samenpflanzen ist der Generationswechsel versteckt, das heißt die Gametophytengeneration lebt nicht selbständig, sondern entwickelt sich auf den Sporophyten und entwickelt sich dort weiter zu den Sporen (Samen).
Blüten
Blüten sind Endständige Sprossabschnitte, die im Dienste der Fortpflanzung ein begrenztes Wachstum haben und im Verlauf dieses Wachstums ihren Vegetationspunkt aufbrauchen (Umsteuerung des Entwicklungsmusters).
Blütenorgane sind Blattmetamorphosen, also modifizierte Blätter. Blütenorgane stehen wirtelig, das heißt die Blütenblätter sind in Kreisen angeordnet, und besitzen keine Internodien. Nodien bezeichnen dabei die Ansatzstellen der Blätter, also sind Internodien die Bereiche der Sprossachse zwischen den Nodien.

Blüte schematisch
Foto: Maksim
Denniss
Rainer_Zenz
Lizenz: [https://www.xonk.de/fdl.txt GNU-Lizenz für freie Dokumentation].
Die Originaldatei ist hier zu finden.
Aufbau einer Blüte
1.Blütenboden
2.Kelchblätter
3.Kronblätter
4.Staubblätter
5.Fruchtblatt oder -blätter
Die Fruchtblätter sind weiter unterteilt in die Narbe(zur Aufnahme des Männlichen Pollens), dem Griffel, durch den die Pollenschläuche zu den Samenanlagen Wachsen und den Samenanlagen, welche im Fruchtknoten (Bauchiger teil der Fruchtblätter) liegen und nach der Befruchtung durch die männlichen Pollen die Samen Bilden.
Der versteckte Generationswechsel der Angiospermen (Bedecktsamer)
Der Generationswechsel der Angiospermen findet in den Blüten statt.
Die Staubblätter bilden dabei die männlichen Microgametophyten innerhalb der Pollen.
Der Fruchtknoten bildet dabei die weiblichen Megagametophyten in Form der Eizelle.
Die Pollen (Microgametophyt)
Pollen entstehen in den Staubblättern. Alle Staubblätter einer Blüte bezeichnet man zusammen als Androeceum.
Jedes Staubblatt besteht aus dem Filament und der Anthere.
Eine Anthere besteht aus 2 Theken (Theka), die wiederum aus je 2 Pollensäcken bestehen.
Ein Pollensack entspricht dabei einem Microsporangium.
Aufbau eines Pollensacks(von außen nach innen)
–Exothecium (Epidermis)
–Endothecium (=Faserschicht)
–Zwischenschicht
–Tapetum (Nährgewebe im jungen Pollensack)
–Archesporgewebe = Pollenmutterzellen (machen die Meiose)
Die Entstehung der Pollen
Die Pollenmutterzellen machen eine Meiose und bilden so die Microsporen, die den jungen Pollen entsprechen, zunächst als Sporentetrade (Gruppen aus je 4). Bei diesem Prozess wird das Tapetum aufgebraucht. Danach zerfallen die Tetraden meist zu Einzelpollen.
Die Mikrosporen machen dann meist noch im Pollensack eine Mitose, wodurch eine vegetative und eine generative Zelle entstehen, welche immer gemeinsam eine Polle bilden. Nun reißen die Theken an einer Sollbruchstelle auf und die reifen Pollen (Microgametophyten) können entweichen.
Aufbau eines Pollens
Die äußere Schicht der Pollen nennt man Exine und ist aus Sporopollenin aufgebaut. Darunter ist eine weitere Schicht, die Intine, welche der Primärwand entspricht.
Die Exine ist besonders interessant, da das Sporopollenin eine besonders hohe chemische Stabilität besitzt und daher die Pollen länger erhalten bleiben können als alle anderen Pflanzenbestandteile. Daraus leitet sich die Palynologie ab, durch die unter anderem durch Analyse fossiler Pollen der einzige Hinweis dafür gefunden wurde, dass Angiospermen bereits in der unteren Kreidezeit existiert haben müssen.
In die Pollenwand sind Öffnungen vorgebildet, sogenannte Aperturen.
Je nach Anordnung der Aperturen auf den Pollen sind die Pollen sulcat (mit einem Öffnungsschlitz), colpat (mit mehereren Öffnungsschlitzen) oder porat (mit porenartigen Öffnungen).
Der Embryosack (Megagametophyt)
Eizellen entstehen in den Fruchtblättern. Alle Fruchtblätter einer Blüte bezeichnet man als Gynoeceum.
Die Fruchtblätter sind mit sich, oder mit anderen Fruchtblättern zum Fruchtknoten verwachsen, welcher je nach Pflanze im Bezug zu den anderen Blütenorganen eine andere Stellung haben kann.
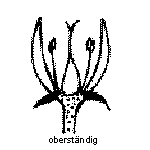
Fruchtknoten Oberständig
Foto: BotBln
Lizenz: [https://www.xonk.de/fdl.txt GNU-Lizenz für freie Dokumentation].
Die Originaldatei ist hier zu finden.

Fruchtknoten Mittelständig
Foto: BotBln
Lizenz: [https://www.xonk.de/fdl.txt GNU-Lizenz für freie Dokumentation].
Die Originaldatei ist hier zu finden.
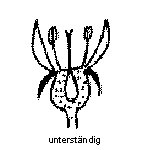
Fruchtknoten Unterständig
Foto: BotBln
Lizenz: [https://www.xonk.de/fdl.txt GNU-Lizenz für freie Dokumentation].
Die Originaldatei ist hier zu finden.
Durch den Fruchtknoten geschützt befinden sich darin die Megasporangien (Nucellus), in dem die Embryosackmutterzelle sitzt, die von den sogenannten Integumenten umgeben sind und über den Funiculus mit dem Fruchtblattrand verbunden sind. Die Öffnung am Ende der Integumente nennt sich Mikropyle.
All diese Struckturen gemeinsam bezeichnet man als Samenanlagen, welche im Bezug zu Funiculus unterschiedlich orientiert sein können und je nach Ausrichtung anatrop(seitlich dem Funiculus angelagert, die Mikropyle zeigt nach unten), atrop(setzt in gerader Linie den Funiculus fort, die Micropyle zeigt nach oben) oder kampylotrop(die Samenanlage liegt seitlich auf dem Funiculus, wobei die Micropyle seitlich nach unten zeigt) genannt werden.
Entstehung des Embryosacks (weiblicher Gametophyt)
Die Embryosackmutterzelle macht eine Meiose, bei der nur 1 Meioseproduckt überlebt. Dieses macht 3 Mitosen, aus denen 8 Zellen entstehen, von denen 2 Zellen zum diploiden sekundären Embryosackkern verschmelzen.
So entsteht der Embryosack (weiblicher Gametophyt), welcher aus 7 Zellen mit insgesamt 8 Zellkernen besteht.
Aufbau des Embryosacks
Am einen Ende des Embryosacks befindet sich die Eizelle welche von den 2 Synergiden umgeben ist.
Am anderen Ende des Embryosacks sind die drei Antipoden.
Zwischen den Antipoden und der Eizelle gemeinsam mit den Synergiden ist Zentral im Embryosack der sekundäre Embryosackkern, welcher das diploide Verschmelzungsprodukt aus den 2 haploiden Zellen ist.
Dies ist der Aufbau des 7 Zelligen, 8 Kernigen Polygonumtyps.
Es gibt auch noch andere Typen, welche auf andere Weise gebildet werden.
Zusammenfassung des Generationswechsels bei Angiospermen
Fruchtknoten (Megasporophyll) —bildet—> Nucellus (Megasporangien) —bildet—> 1Überlebendes Meioseprodukt (Megasporen) —bildet—> Embryosack (Megagametophyt) mit Eizelle (Gamet)
Staubblatt (Mikrosporophyll) —bildet—> Pollensack (Mikrosporangien) —bildet—> Junges Pollenkorn (Mikrosporen) —bildet—> Pollen (Mikrogametophyt)
[printfriendly]
hallo ich habe einen kleines Staubblatt,
liebe grüsse,
felipe schulz