
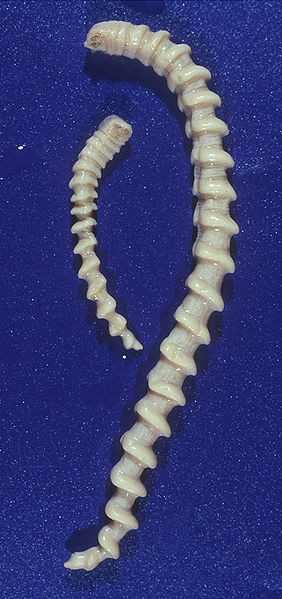
männliche (links) und weibliche (rechts) Adulten der Armillifer armillatus
Foto: Dennis Tappe & Dietrich W. Büttner
Lizenz: Creative Commons-Lizenz Namensnennung 2.5 US-amerikanisch (nicht portiert)
Die Originaldatei ist hier zu finden.
Einleitung
Die Suche nach der Herkunft der rein parasitisch lebenden Zungenwürmer erweist sich als kontrovers. Im Laufe der Geschichte der Zoologie wurden sie mit den folgenden Gruppen verglichen: Tardigrada, Olychophora und Annelida, mit den Arthropoda im allgemeinen und mit den Milben,Insekten, Krebsen und Tausendfüßlern.
Wie viele parasitisch lebende Tiere (z.B. Copepoda, Cirripedia, Isopoda) hat diese Lebensweise tiefgreifende Veränderungen im Körperbau der Zungenwürmer mit sich gebracht, sodass es Systematikern schwer fällt, sie einzuordnen. Die extreme Reduktion, die mit der parasitischen Lebensweise einher geht, lässt kaum noch Rückschlüsse auf ihre Herkunft zu. Während die Larven anderer Parasiten es noch erlauben, die Tiere in einem System zu erfassen, versagt auch diese Methode bei den Larven der Zungenwürmer.
Neben den morphologischen Untersuchungen unterstützen molekularbiologische Methoden zunehmend die Hypothese, welche die Zungenwürmer bei den Crustaceen einordnet. Dagegen sprechen einige mutmaßliche marine Vertreter der Zungenwürmer aus dem späten Kambrium.
Die Klassifizierung in diesem Artikel folgt der von Martin und Davis (2001):
Stamm: Arthropoda
Unterstamm: Crustacea
Klasse: Maxillopoda
Unterklasse: Pentastomida
Ordnung: Cephalobaenida
Familie: Cephalobaenidae
Reighardiidae
Ordnung: Porocephalida
Familie: Armilliferidae
Diesingidae
Porocephalidae
Sambonidae
Sebekiidae
Subtriquetridae
Linguatulidae
Lebensweise
Derzeit sind über 130 Arten der Unterklasse Pentastomida bekannt, von denen alle obligate Parasiten sind, d.h. sie brauchen für ihre Entwicklung zwingend einen Wirt. Zu den Wirten zählen überwiegend Reptilien (90%), hundeartige Raubtiere und Vögel. Die Entwicklungsstadien der Zungenwürmer finden sich in den unterschiedlichsten Organen aller Wirbeltiere, aber auch in denen einiger weniger Insektenarten.
Die adulten Formen leben überwiegend in den Atemorganen (Lunge), Atemwegen (Nasengänge) und ihren Nebenräumen (Luftsäcke) der landlebenden, fleischfressenden Vertebraten und saugen nach dem Anheften an den Lungenkapillaren Blut, oder ernähren sich, wenn sie sich in den Nasengängen befinden, von Nasenschleim mit den darin befindlichen abgestoßenen Epithelzellen und von Lymphe.
Die genaue Ernährungsweise und die physiologischen Vorgänge bei der Verdauung der Pentastomida sind noch weitestgehend unklar.
Bei einer Studie (1973) mit Reighardia sternae konnte Riley ansatzweise der Blutaufnahme auf die Spur kommen. Nach dem Aufspüren einer Lungenkapillare klammert sich Reighardia sternae an ihr fest und zieht die Kapillarwand mit der Pharynxmuskulatur blasenförmig in die Mundhöhle, in der sie durch eine muskulöse Oralpapille zum Platzen gebracht wird.
Weiterhin konnte beobachtet werden, dass weibliche Zungenwürmer kaum ihren einmal gefundenen Aufenthaltsort verlassen, männliche Zungenwürmer hingegen öfter.
Anatomie
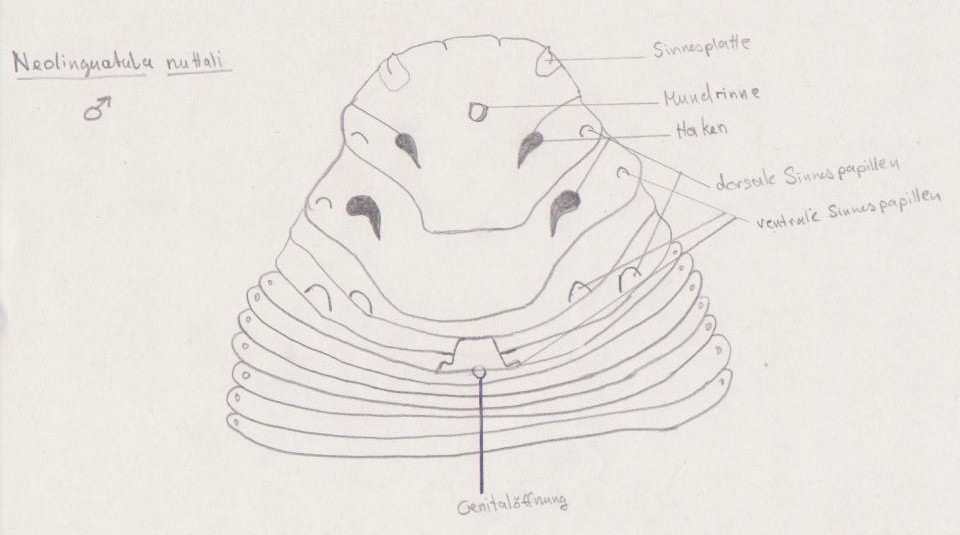
männliche Neolinguatula nuttali Ventralseite des Vorderkörpers
Die Größe der Arten variiert zwischen 1cm – 16cm, mit den weiblichen Armillifer armillatus als größte Art (Abb.1). Die die Nasenhöhlen bewohnenden Zungenwürmer sind meist dorsoventral abgeflacht, die Lungen bewohnenden dagegen im Querschnitt trapezförmig bis rund. Ihr Körper kann in zwei Regionen unterteilt werden: Der vergleichsweise kurze Kopf und der lange geringelte Rumpf.
Die Bezeichnung „Fünfmünder“ ist irreführend. Der Kopf besitzt neben der Mundöffnung 2 Paar Klammerhaken aus Chitin, welche aus den Hauttaschen herausgestreckt werden können. Bei primitiven Formen (Ordnung Chephalobaenida) befinden sich die Haken an der Spitze von stummelartigen Auswüchsen. Durch Flüssigkeitsdruck können sie gestreckt und durch Muskelzug wieder gekürzt werden.
Ebenfalls zur Kopfregion zählen die Apical- und Frontalpapillen (Abb. 2). An das Vorderende schließt sich der häufig geringelte Rumpf. Die beiden Abschnitte gehen meist ohne scharfe Grenze ineinander über (Abb. 1), doch es existieren auch Arten, bei denen eine deutliche Einschnürung zu erkennen ist (Cubirea annulata), welche Rückschlüsse auf die jeweilige Spezialisierung an den Lebensraum einer Art erlaubt. Die Anzahl der Rumpfringe variiert zwischen den männlichen und weiblichen Individuen, kann aber auch bei den höher entwickelten Porocephalida deutlich größer sein als bei den Cephalobaenida. Die äußere Ringelung ist ein mögliches Indiz für eine echte aber unvollständige Metamerie.
Integument und Muskulatur
Das Integument der Pentastomida besteht aus einer chitinhaltigen Kutikula, die von den Zellen der einschichtigen Epidermis abgesondert wird. Die Kutikula, an deren Oberfläche die Kanäle der Drüsenzellen enden, unterteilt sich von oben nach unten Richtung Epidermis in Epi-, Exo und Endokutikula. Während die Cephalobaenida einzellige Stigmendrüsen besitzen, die überwiegend auf das Vorderende, besonders im Umkreis der Klammerhaken, verteilt sind, besitzen die höher entwickelten Porocephalida mehrzellige Stigmendrüsen, die auf dem ganzen Körper zu finden sind.
Unter dem Integument kommt der Hautmuskelschlauch mit der dünnen äußeren Ringmuskulatur und der stärkeren Längsmuskulatur, die in das Bindegewebe eingelagert ist. Es handelt sich ausschließlich um quergestreifte Muskulatur.
Nervensystem
Die höher entwickelten, ausgewachsenen Porocephalida besitzen ein Nervensystem, das stärker reduziert ist als das der Cephalobaenida. Das Nervensystem der Cephalobaenida hat eine zum Teil noch äußerlich erkennbare metamere Anordnung der Ganglienpaare, das der Porocephalida besteht aus einem einzigen ungegliederten postoralen Nervenknoten und einer praeoralen Komissur ohne Ganglienzellen.
Wie vielen Parasiten fehlen den Zungenwürmern komplizierte Sinnesorgane wie Augen, stattdessen besitzen sie als Sinnesorgane Chemo- und Mechanorezeptororgane in Form von Apical-, Frontal- und Lateralpapillen (Abb. 2). Diese Sinnespapillen können durch Änderung der Turgordrucks ausgestreckt und durch Muskeln eingezogen werden.
Verdauungstrakt
Der Verdauungstrakt ist auf das wesentlichste reduziert:
Ein nahe dem Vorderende liegender ventraler Mund, ein Vorderdarm bestehend aus der Mundhöhle, dem Pharynx, der als Saugpumpe dient, und dem Oesophagus. Der sehr einfach gebaute Mitteldarm und der Enddarmbereich besitzt keine Ausstülpungen und Anhangsdrüsen. Nur Vorder- und Enddarm sind mit einer Chitinintima ausgekleidet.
Atmungsorgane, Kreislauforgane oder besondere Exkretionsorgane fehlen. Es wird vermutet, dass die Stigmendrüsen bei der Exkretion eine Rolle spielen.
Geschlechtsorgane
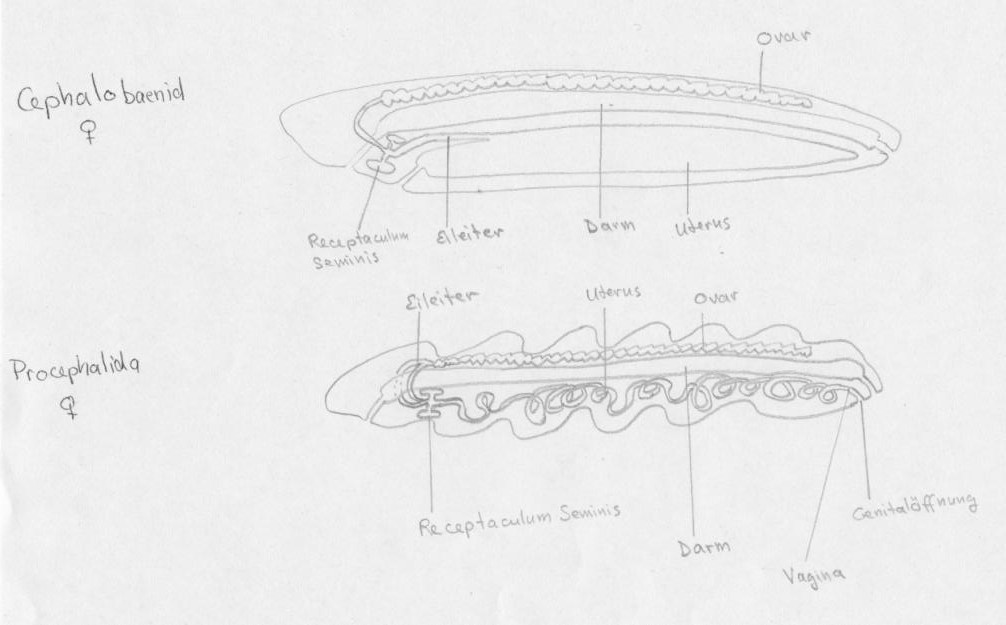
weibliche Cephalobaenid und Procephalida
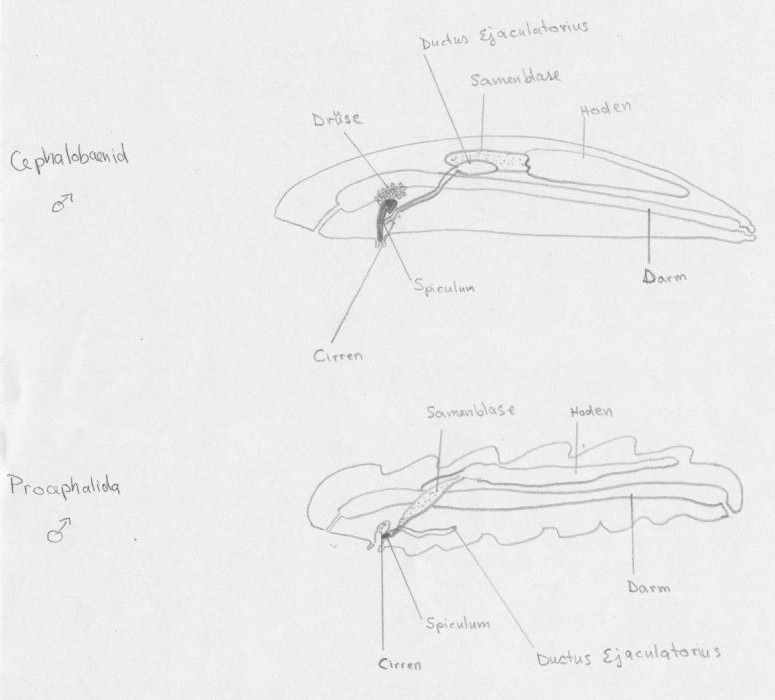
männliche Cephalobaenid und Procephalida
Zungenwürmer sind getrenntgeschlechtlich.
Weibliche Zungenwürmer besitzen meist ein dorsales, blind endendes und schlauchförmiges Ovar. Während die Cephalobaenida (Abb. 3) nur einen Eileiter besitzen, haben die Porocephalida (Abb. 3) zwei Eileiter.
Bei den Cephalobaenida führt der Eileiter zu den zwei Spermavorrattaschen (Receptaculum seminis), gefolgt von dem sackförmigen Uterus, welcher durch die Vagina mit der Genitalöffnung in Verbindung steht.
Die zwei Eileiter der Porocephalida schließen sich unterhalb des Darmes zusammen und an diesem Punkt sind zwei Spermavorrattaschen durch kleine Kanälchen angelagert. Der schlauchförmige Uterus windet sich mehrere Male im weiblichen Körper und endet in die posterior-gelegene Genitalöffnung. Die Lumen der Vagina und des Uterus sind in beiden Ordnungen von Chitinintima ausgekleidet.
Alle männliche Zungenwürmer besitzen einen dorsalen, unpaarigen Hoden (Abb. 4), nur Linguatula spp. besitzen zwei. Bei Cephalobaenida ist der Hoden durch einen muskulösen Schleusenmechanismus von der Samenblase abgetrennt, der sicher stellt, dass nur reife Samen in die Samenblase gelangen. Von der Samenblase führen zwei Kanälchen zu dem muskulösen Spritzkanal (Ductus ejaculatorius). Die Vasa deferentia schließt sich an und mündet in dem paarigen Begattungsapparat.
Studien zeigen, dass die Paarung nur zu einem bestimmten Zeitpunkt in dem Endwirt stattfinden kann, nämlich wenn die Weibchen noch jung sind und die Vagina und die Spermavorrattaschen nahe genug für die Spermieninjektion durch die Cirren sind. Die Spermien wandern durch das Lumen zum Rand des Ovars, wo sie durch eine Basallamina am Weiterwandern gehindert werden.
Nach der Entwicklung der Eier, welche erst später nach der Imaginalhäutung stattfindet, wandert die Eizelle durch die Eierstockwand und wird zum Eileiter transportiert. Nach der Befruchtung wird die Vagina zum Uterus, welcher so stark in die Länge wachsen kann, dass die älteren Weibchen nicht mehr begattet werden können. Nach der Befruchtung erreichen die Eier den Uterus, in dem die Entwicklung zum ersten Larvenstadium erfolgt.
Entwicklung
In dem Uterus entwickeln sich die Eier unterschiedlich schnell, von daher kann man alle Stadien der Entwicklung bis zum 1. Larvenstadium im Uterus vorfinden.
Es zeigte sich, dass das Muttertier sich erst öffnet, wenn mindestens 25% aller Eier sich vollständig zum Embryo entwickelt haben.
Die primäre Larve ist die erste ansteckende Phase für den nächsten Wirt. Abhängig von der Art kann der Zwischenwirt ein Invertebrat oder ein Vertebrat sein. Ob ein Zwischenwirt fakultativ oder obligatorisch ist, ist von der Art abhängig.
Wird ein Zwischenwirt oral befallen, gelangt die primäre Larve über den Verdauungstrakt in die Leibeshöhle und von dort zu den verschiedensten Organen. Dort verkapselt sich die Larve und entwickelt sich so weit, dass sie infektiös für den Endwirt ist.
Die Zahl der Nymphenstadien der Zungenwürmer variiert. Anhand von Raillietiella spp. wurde gezeigt, dass beim Befall von Geckos die infektiöse Phase das 3. Nymphenstadium (d.h. nach zwei Häutungen) ist, aber bei anderen Arten erst das 4. Nympfenstadium die infektiöse Phase darstellt.
Generell kann gesagt werden, dass bei Vertebraten als Zwischenwirt 4 Häutungen von Nöten sind, einige Porocephalida jedoch mehr Häutungen (6-8) benötigen.
Nimmt der Endwirt den Zwischenwirt oder einen Teil von ihm in sich auf, gelangt die infektiöse Larve über den Verdauungstrakt in die Atemorgane (Lunge), die Atemwege (Nasengänge) und in ihre Nebenräume (Luftsäcke).
Um reif für die Fortpflanzung zu werden, benötigen die Zungenwürmer wiederum mehrere Häutungen, deren Zahl von Art und Ordnung abhängt.
Die Anzahl der produzierten Eier kann unterschiedlich hoch sein: Während Reighardia spp. ungefähr 3.000 Eier produziert, sind es bei Linguatula serrata mehrere Millionen Eier.
Zusammenfassung
Obwohl die Pentastomida aufgrund molekularbiologischer Untersuchungen heute bei den Crustaceen eingeordnet werden, bleiben aufgrund verschiedener morphologischer Untersuchungen sowohl an den Spermien als auch an den folgenden Stadien und den Fossilfunden weiterhin Zweifel an dieser Einordnung bestehen.
Die phylogenetische Position der Parasiten allein durch morphologische Daten zu bestimmen, ist fast unmöglich. Viele Parasiten haben sich in ihrer Entwicklung und Morphologie stark verändert, um sich an die neue Lebensweise im Wirt anpassen zu können. Bei den Pentastomida war diese Anpassung so umfassend, dass die Reduktion auf die grundlegenden Körperfunktionen die Einordnung extrem erschwert. Die Cuticula und ihre Struktur, die kurzen, gegliederten Anhänge mit den Krallen, und die Ähnlichkeit der Spermien und deren Entwicklung mit den Spermien von Fischläusen und deren Entwicklung deuten auf eine Verwandtschaft der Pentastomida mit den Arthropoden hin.
Dennoch fehlen ihnen grundlegende Eigenschaften der Arthropoden wie eine klare Körpersegmentierung, Nephridien, das Blutkreislaufsystem und die typische Gehirnentwicklung. Aus diesem Grunde und aufgrund von Fossilfunden, die aus dem späten Kambrium stammen, ordnen einige die Pentastomida außerhalb der Crustaceen ein und einige sogar außerhalb der Arthropoden.
Sollten die molekularen Untersuchungen allerdings stimmen, würde das im Fall der Zungenwürmer bedeuten, dass sie viele wichtige phylogenetische Eigenschaften verloren haben, die ihre Position bei den Arthropoden und speziell den Crustaceen untermauern könnten.
[printfriendly]